原孙等南京林业大学学报


专题报告
基于CRISPR/Cas9 的毛白杨转录因子bHLH106功能研究
孙、郭燕娇、、周*、姜立全、李伟*
林业遗传育种国家重点实验室(东北林业大学)

木材形成机理的研究一直是林业研究的重点。木材的形成是一个高度有序和连续的过程,包括维管形成层的增殖、木质部细胞的分化和膨胀、次生细胞壁的沉积和加厚以及细胞的程序性死亡。次生木质部包括纤维细胞、导管分子细胞和射线细胞。许多参与次生木质部发育的转录因子已经从模式植物中被鉴定出来,它们形成一个转录调控网络来调控木材的形成过程。因此,探索转录因子在木材形成过程中的作用机制,可以为利用树木分子生物学手段改良木材性质、提高木材质量提供理论依据。
BHLH(碱性螺旋-环-螺旋)是一种碱性螺旋-环-螺旋转录调控因子,主要存在于真核生物中,是仅次于MYB的第二大转录因子家族。在植物中,bHLH转录因子家族在调节植物的发育、胁迫反应、信号转导和二次生长中起着重要的作用。大多数bHLH蛋白是在模式植物拟南芥中发现和鉴定的。它们的功能包括:参与信号转导合成代谢途径的调节,如植物激素的合成、光信号转导、光敏信号调节和脱落酸信号转导;参与植物的生长发育,如种子萌发;植物抗逆性的调节,如低温胁迫、盐胁迫等。此外,bHLH基因家族还参与木质部的生长发育,例如,一个非典型的bHLH转录因子调控生长素下游的早期木质部发育;棉花bHLH蛋白GhFP1正向调节纤维细胞的伸长;在拟南芥次生细胞壁的转录组数据中,发现了一些bHLH转录因子调控次生细胞壁的合成。在一项研究中,确定了TMO5/LHW(一种bHLH转录因子二聚体)是拟南芥中导管发育的关键调节因子,并且在其缺失突变体中导管组织逐渐消失。BHLH转录因子LHW-T5L1二聚体促进木质部前体细胞中关键细胞分裂素基因的表达,导致周围细胞中细胞分裂素水平升高,从而促进原形成层细胞的特异性增殖。然而,很少报道bHLH转录因子参与木材形成。
基因编辑工具的出现为基因功能的研究和植物性状的改良带来了巨大的帮助。Crispr(聚类规则间隔短回文重复序列)基因编辑技术极大地提高了基因编辑的效率和实用性。目前,该技术已广泛应用于基础科学、人类疾病治疗、作物基因功能、遗传育种等领域。CRISPR/Cas9系统由两部分组成:指导RNA(sgRNA)和Cas9核酸酶。该系统的主要功能是在特定的基因组位点诱导DNA双链断裂。双链断裂后非同源末端连接的细胞修复可能导致碱基的插入或缺失,从而改变碱基序列。CRISPR/Cas9技术可用于编辑全基因组中的任意基因,通过CRISPR/Cas9系统可获得产量、品质和抗性更高的优良品种。比如通过CRISPR/Cas9技术编辑马铃薯R基因,可以获得抗病的马铃薯品系。通过CRISPR介导的CCR基因编辑技术获得了优良的低木质素杨树新品系。自2015年CRISPR/Cas9系统首次应用于杨树以来,已有许多研究将CRISPR/Cas9系统应用于树木的基因功能。例如,CRISPR/Cas9技术被用于创建毛白杨PtrVCS2基因的突变体。通过对突变植株的分析,发现PtrVCS2促进形成层分化为木质部,使植株提前产生加厚的次生细胞壁,说明毛白杨PtrVCS2参与了木材形成的调控。利用CRISPR/Cas9技术,获得了毛白杨形成层特异表达的PtrWOX4转录因子功能缺失突变体,该突变体破坏了植株的顶端优势,使顶芽和根系发育不良,显著影响形成层发育,木质部分化紊乱。
东北林业大学林木遗传育种国家重点实验室的姜立权通过分析野生型毛白杨茎不同细胞类型的RNA-seq数据,鉴定出一个在形成层和次生木质部细胞中高表达的PtrbHLH106基因,并利用CRISPR/Cas9系统构建了毛白杨ptrbhlh 106基因的功能缺失突变体。通过对突变体的表型观察和分析,初步揭示了该基因在木材形成中的功能。
跟期刊君一起来了解一下吧!
作者简介
联系人

周,女,1986年9月生,东北林业大学教师,主要从事木质素生物合成调控和林木基因编辑研究。
联系人

李伟,男,1983年5月出生,本刊青年编辑,东北林业大学林木遗传育种学科教授、博士生导师。主要从事林木遗传改良的分子基础研究,重点研究木材形成调控的分子机制、林木次生生长和环境适应机制。
第一作者

孙,女,1997年3月出生,东北林业大学林木遗传育种专业研究生。

关键词:毛白杨;次生木质部发育;CRISPR/Cas9基因编辑;PtrbHLH106转录因子
基金项目:国家自然科学基金委青年科学基金项目(32001332);中央高校基本科研业务费专项资金项目(2572018CL01)。
引用格式:孙,郭燕娇,,等.基于CRISPR/Cas9的毛白杨转录因子bHILH106的功能研究[J].南京林业大学学报(自然科学版),2021,45 (6): 15-23。孙君太。等.基于CRISPR/Cas9的毛果杨bHLH106转录因子的功能研究[J].南京林业大学学报(自然科学版),2021,45(6):15-23。DOI:10.12302/j ISSN . 1000-2006.2107031。


1目的
利用CRISPR/Cas9基因编辑系统构建了毛果杨bhlh06(碱性螺旋-环-螺旋106)基因突变体,并对其表型特征进行了分析,初步揭示了PtrbHLH106基因在毛果杨木材形成过程中的功能。
2方法
基于野生型毛白杨茎不同细胞类型(形成层、木质部和韧皮部细胞)的RNA-seq数据,克隆了一个在形成层和木质部高表达的bHLH基因PtrbHLH106。利用CRISPR/Cas9基因编辑技术构建毛白杨PtrbHLH106功能缺失突变体。对生长60、90和120天的毛白杨ptrblh106突变体和野生型植株进行了表型观察。在120天的植物的每个茎节上制作石蜡切片,通过甲苯胺蓝染色观察并通过细胞统计分析。
2.1测试材料
实验材料为东北林业大学林木遗传育种国家重点实验室(本实验室)保存的毛白杨植株,基因型为Nisqually-1。用次氯酸钠对野生毛白杨侧枝消毒后,通过植物组织培养诱导出芽,获得毛白杨无菌组培苗。组织培养条件:温度25℃,光强40 μ mol/(m2.s),光周期16 h光照/8 h黑暗。温室条件:温度范围22~25℃,光照强度约300 μ mol/(m2.s),光周期16 h光照/8 h黑暗。温室栽培毛白杨,采用135℃灭菌30分钟后冷却的土壤。
PENTR载体(pENTR/D-topo)购自Life In-vitrogen公司;T载体(pMD18-T)购自TaKaRapUC19-GFP载体保存在我们的实验室中。PEgP237-2A-GFP由Keishi Osakabe实验室捐赠。RNA试剂盒(RNease plant Mini Kit)、凝胶提取试剂盒、PCR纯化试剂盒、质粒Mini Kit均购自QIAGEN公司。逆转录试剂盒(PrimeScript RT试剂盒)购自TAKARAPfu聚合酶(Pfu DNA聚合酶)购自Aglient公司;体外转录试剂盒(T7快速高产量RNA合成试剂盒)购自NEB甲苯胺蓝染色剂购自适马;大肠杆菌)TOP10和根癌农杆菌GV 3101的感受态细胞均保存在本实验室。
2.2研究方法
2.2.1建立层级模型
根据PtrbHLH106基因(potri.006g037100)的id,从毛白杨基因组网站(https://phyto zome . jgi . doe . gov/pz/portal . html)下载基因序列,设计特异性引物(表1)。利用RNA提取试剂盒从毛白杨分化木质部中提取总RNA。用逆转录试剂盒逆转录获得的cDNA作为PCR扩增模板,扩增目的基因。PCR反应体系为10×Pfu缓冲液2 μL,dNTPs 2 μL,cDNA模板1 μL,正向和反向引物1 μ L,Pfu DNA聚合酶0.4 μL,去离子水12.6 μL,反应体系为20μL . PCR反应程序:94℃预变性2min;94℃变性45 s,52℃退火30 s,68℃延伸30 s,30个循环;68℃再加热30分钟。通过PCR扩增获得基因片段,并连接到pENTR载体上。经热休克大肠杆菌转化后,获得阳性克隆,测序检测PtrbHLH106序列。
▼表1使用的引物和序列

3个结果
获得毛白杨ptrbhlh106突变体;与野生型相比,突变体植株的株高和地径没有显著差异。在整个测定的生长周期中,第8茎节的长度有缩短的趋势,茎节的数量有增加的趋势。形成层细胞的数量有增加的趋势,但差异不显著。导管分子的孔径明显增大,成纤维细胞的数量明显减少。
3.1 获得ptrbhlh106 基因
研究团队通过激光捕获显微切割(LCM)精确采集毛白杨野生型植株第8茎节的形成层、木质部和韧皮部细胞,并通过RNA-seq数据进行分析。结果表明,PtrbHLH106基因主要在形成层和木质部细胞中表达(图1),因此推测PrbHLH106可能在形成层和木质部发育中起一定的调控作用。通过克隆和测序PtrbHLH106,获得了723 bp的CDS序列。
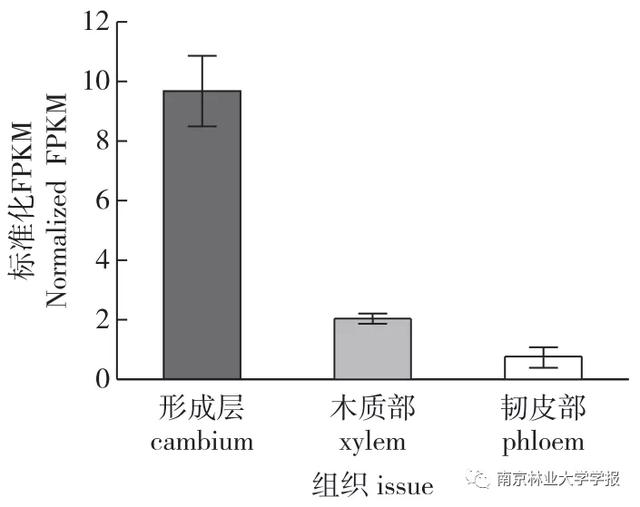
▲图1 ptrbhlh 106基因在毛白杨不同组织细胞类型中的表达
注:标准化FPKM值是指转录本丰度被标准化为每百万个图谱中每千个转录碱基读取的片段数,误差线代表三个生物重复的标准误差。
3.2毛白杨ptrbhlh 106基因突变体的创建
为了进一步研究PtrbHLH106在木材形成过程中的功能,利用CRISPR/Cas9基因编辑技术构建了毛白杨PtrbHLH106基因功能缺失突变体。在创建突变体之前,通过体外切割技术验证所选的gRNA,以确定gRNA的可用性。
3.2.1体外验证GRNA
根据gRNA网站的预测,选择了一个得分高、缺失概率低、GC含量约为50%的PrbHLH106外显子106上的gRNA。切割模板的示意图如图2A所示,gRNA位于线性化DNA模板的-952 bp处。体外切割后,琼脂糖凝胶电泳的结果如图2B所示。当不加入gRNA或Cas9蛋白时,含有PtrbHLH106基因片段的模板DNA不被切割(泳道1和2)。加入gRNA、Cas9蛋白和模板DNA后,DNA被有效地切割成2段,电泳条带约为4.5 kb和0.9 kb(泳道3),与理论长度一致。结果表明,所选的gRNA序列能有效结合Cas9蛋白并切割靶位点,可用于后续实验。
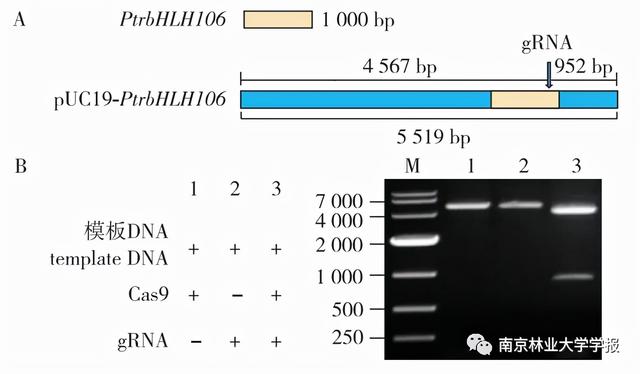
▲图2 GRNA的体外验证
注:a .模板DNA示意图;b .凝胶电泳结果;1.2.对照组;3.测试样本。
3.2.2毛白杨的遗传转化及转基因植株的鉴定
构建了CRISPR载体pegp 237-U6-ptrbhlh 106 grna-35S-cas 9,通过农杆菌感染、抗性愈伤组织萌发和抗性芽在卡那霉素培养基上两次生根筛选的过程,获得了三株抗性生根植株(图3a-3c)。提取待鉴定的抗性植物的叶的基因组DNA,并通过使用载体引物M13-F和PtrbHLH106的gRNA-R末端引物进行PCR扩增。如图3D所示,两个阴性对照(泳道1和3)没有扩增出片段条带,而抗性植物(泳道4至6)都扩增出与阳性对照(泳道1)相同大小的条带,这表明ptrbhlh100。

▲图3毛白杨转基因植株的获得
注:a .愈伤组织分化诱导;b .诱导抗性芽;筛选抗性植物;转基因植物的PCR鉴定。1.DDHO是模板的阴性对照;2.以pegp 237-U6-ptrbhlh 106 grna-35S-cas 9质粒为模板的阳性对照;3.以野生毛白杨叶片DNA为模板的阴性对照;4~6.以抗性植物叶片的DNA为模板的样品。
3 . 2 . 3ptrbhlh 106的基因编辑分析
通过测序鉴定转基因植物中PtrbHLH106基因的靶位点的编辑。结果如图4A所示:三个菌株的突变gRNA中插入或缺失了不同数量的碱基,如在ptrbhlh106-1的两个等位基因中分别插入了1个碱基和2个碱基,在ptrbhlh106-2的两个等位基因中分别插入了1个碱基和3个碱基,在ptrbhlh 106-3的两个等位基因中分别插入了1个碱基和3个碱基。三个突变植株都是双等位基因突变。编辑PtrbHLH106基因后的蛋白质翻译如图4B所示。插入和删除一些碱基后,翻译提前终止,产生的氨基酸序列比正常的ptrbHLH106短得多。ptrbhlh106-2的一个等位基因缺失一个氨基酸也引起一个氨基酸的改变。其他基因序列的翻译序列不含bhlh家族典型的HLH结构域。结果表明,成功获得了毛白杨ptrbhlh106突变体。为了验证PtrbHLH106转录因子的功能,选择突变植物ptrbhlh106-2、ptrbhlh106-3-3进行后续研究。
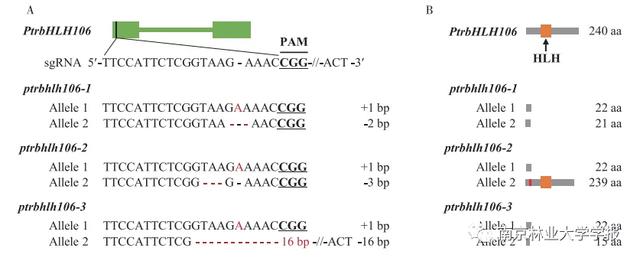
▲图4毛白杨PtrbHLH106基因的编辑
注意:a .等位基因编辑(红色字母表示碱基插入,红色& # 34;-"表示碱基缺失,& # 34;-16 bp-“表示缺失16个碱基);b .编辑基因的蛋白质翻译预测(红色块区域代表氨基酸变化)。
3.3毛白杨ptrbhlh 106突变体的表型分析
ptrbhlh106突变株无性繁殖后,与高度相似的野生毛白杨组培苗一起盆栽于温室,在植株自然生长60、90、120天时进行表型观察(图5)。从生长过程来看,毛白杨ptrbhlh106突变体的株高和地径与野生型没有显著差异。同时测量茎节数和第8茎节的长度。统计结果表明,生长60天时,突变体植株的茎节数显著大于野生型。随着植株生长时间的延长,茎节数有增加的趋势,但差异不显著。在植物生长的第60天和第120天,第8个茎节的长度与WT显著不同。从植物生长周期来看,突变株的第8茎节长度有缩短的趋势。从以上结果初步推测PtrbHLH106功能缺失,对植物生长无严重影响。
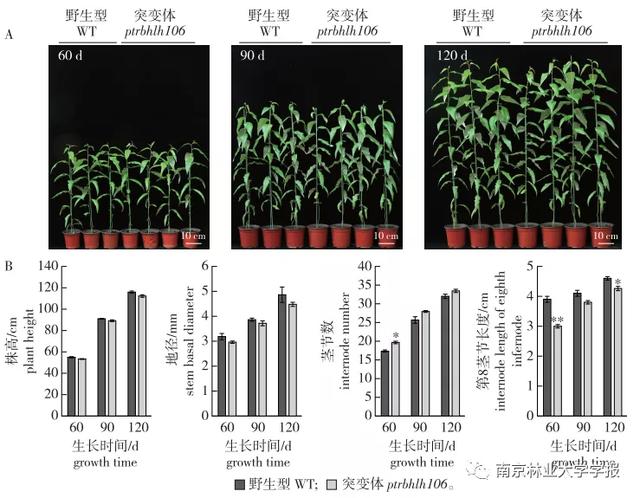
▲图5毛白杨野生型和ptrbhlh106突变体的表型分析
注:误差线代表通过三次生物重复计算的标准误差,*表示通过1检验,突变体和野生型植物株系之间存在显著差异,*。P
为了进一步分析PtrbHLH106在杨树木材形成过程中的作用,对毛白杨突变体bHLH106的第2、4、6、8、10茎节横切面进行石蜡切片观察,结果表明突变体的各类细胞形态与野生毛白杨无差异(图6A)。切片中导管分子、单位面积纤维细胞数和单个导管分子孔径的统计分析表明,与野生型相比,突变体植株的纤维细胞数显著减少,导管分子数差异不显著,单个导管的孔径显著增大。通过观察和分析形成层细胞,发现突变体植株的形成层有增加的趋势,但差异不显著(图6B和6E)。以上结果表明PtrbHLH106转录因子功能缺失导致木质部纤维细胞减少,导管直径增大,初步表明该转录因子在次生木质部发育中起调控作用。

▲图6毛白杨ptrbhlh106突变体石蜡切片观察和细胞统计
注:a .野生型(WT)和突变体(ptrbhlh106)毛白杨茎节细胞形态观察(比例尺:200微米);b .观察形成层细胞的形态(红线表示形成层面积,尺度为100微米);c .计算每单位面积的细胞数;d .统计导管的直径和面积;计算形成层细胞的数量。
4结论
ptrlh106突变体与野生型植株在导管直径和纤维细胞数量上存在差异,初步证明ptrbhlh106基因参与调控毛白杨次生木质部的发育。

关注我们,更多精彩